
|
|
Biología del Virus de la Polio.
Breve
Resumen
Por
|
Clasificación Familia
Picornaviridae, género enterovirus, especie virus polio (VP).
Protege al genoma viral y da la estructura a la partícula vírica. Está formada por cuatro proteínas estructurales, denominadas VP1, VP2, VP3 y VP4. Las tres primeras son externas y la cuarta es interna y está unida covalentemente al extremo 5´ del genoma viral. En la superficie del virión (Figura 1) hay unas zonas que sobresalen, y son las encargadas de activar la respuesta inmune; zonas donde se unirán los anticuerpos específicos neutralizantes y otras zonas deprimidas, por donde el virus interacciona con el receptor celular, que es una inmunoglobulina (Ig) del sistema nervioso central, para infectar a las células diana (las neuronas). El ciclo de la replicación
viral Los virus son incapaces de reproducirse por sí solos. Necesitan utilizar parte de la maquinaria biosintética de las células específicas que infectan, en detrimento de éstas, por lo que se les llama parásitos celulares.
Los
virus tienen una apetencia específica por determinados huéspedes. Esta
característica se llama tropismo celular. Así, por ejemplo, el VP sólo
infecta al hombre y, de forma experimental, también a algunos primates,
pero no infecta a otros animales. Además, no todos los órganos, ni
cualquier célula le sirven, sólo aquellas que tengan el receptor adecuado,
convirtiéndolas en células susceptibles. Para el VP, éstas son las células
del sistema nervioso central. Cuando el VP entra en contacto con la célula
adecuada, se une a los receptores celulares quedando adsorbido a la
membrana celular. Ésta le rodea completamente formando una vesícula
llamada endosoma, que le transporta al interior de la célula; al
citoplasma. Dentro del endosoma, unas enzimas celulares rompen la
nucleocápside (proteasas) y liberan el ARN viral al citoplasma. Este se
introduce en un ribosoma celular, desde donde inicia su replicación,
sintetizando los componentes necesarios para formar viriones nuevos; es
decir, la nueva progenie viral (Figura 2). Esto provoca una limitación o,
incluso la inhibición del metabolismo celular, ya que algunas proteínas
celulares van a ser utilizadas por el virus, con el consiguiente perjuicio
para la célula. El ARN
viral tiene capacidad para replicarse y para funcionar como ARN mensajero,
traduciéndose a la gran proteína, llamada poliproteína, utilizando la mayoría del genoma
(aproximadamente el 90%). Esta poliproteína es el precursor de las
proteínas víricas. La poliproteína inmediatamente se rompe, por acción de
las proteasas, y da lugar a proteínas más pequeñas; cuatro estructurales
que forman la nucleocápside y otras que no son estructurales, y no
forman parte del virión, pero son necesarias para la replicación viral y
para inhibir la síntesis de componentes celulares. Entre las proteínas no
estructurales está la ARN polimerasa vírica, que será la encargada de
dirigir la síntesis de las cadenas nuevas del genoma viral, el VP la
sintetiza porque en la célula no existe ninguna enzima con esa
actividad. Por
acción de la ARN polimerasa vírica, cada molécula del ARN positivo vírico
se copia a una cadena complementaria de ARN negativo, que será utilizado
como molde para volver a ser copiado a una cadena nueva de ARN positivo. De esta forma, la
información genética es copiada íntegramente originando cadenas casi
idénticas nuevas del genoma viral original. Este ciclo se repite muchas
veces, mientras que la infección esté activa. Con
las proteínas estructurales recién sintetizadas, se forma la
nucleocápside. Después entra el ARN nuevo, plegándose para adaptarse al
hueco y de esta manera, se forma el virión nuevo. Los virus nuevos se
acumulan en el citoplasma hasta que la célula se rompe y son liberados al
exterior como partículas infecciosas con capacidad para infectar células
próximas y repetir el ciclo hasta que la infección se para por acción de
los anticuerpos neutralizantes,
que el organismo huésped sintetiza para abortar la
infección. 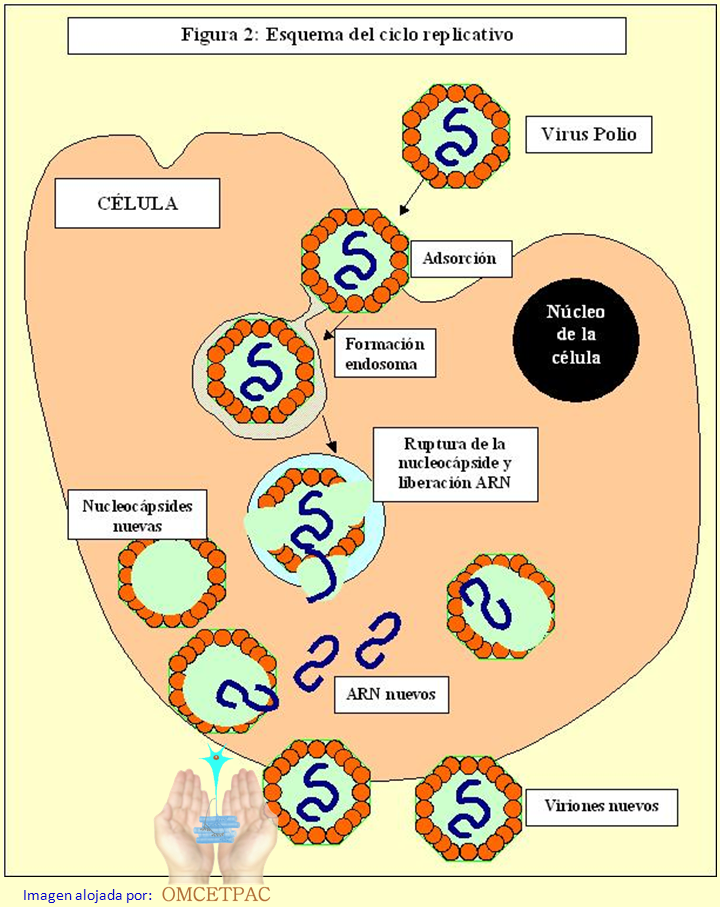 Durante un tiempo, llamado de eclipse, no se detectan viriones infecciosos dentro de la célula. Esta fase se corresponde con la ruptura de la nucleocápsida y la liberación del ARN viral, hasta la formación de los nuevos viriones. La replicación viral no siempre ocurre con la misma eficacia. No todos los virus infecciosos se adsorben a la célula. De los que se adsorben, algunos no penetran en el interior celular. Se sintetiza mucho material vírico, proteínas y ARN, pero no todo se utiliza para forman viriones completos, pueden quedar nucleocápsidas vacías, son las llamadas partículas defectivas que no son infecciosas. La replicación viral cesa cuando actúan los anticuerpos neutralizantes específicos. Estos anticuerpos sintetizados por los linfocitos B, que han sido estimulados previamente por las proteínas externas de la nucleocápside (antígenos), se unen a las zonas prominentes del virión, lo rodean y ocultan las zonas deprimidas donde se tendrían que unir los receptores celulares para iniciar la infección. A medida que la infección progresa, aumenta la síntesis de anticuerpos, hasta que estos prevalecen, neutralizando todos los viriones nuevos, abortando y resolviendo la infección. Variabilidad
genética Los virus ARN poseen una variabilidad genética. Esto ocurre porque durante la replicación del ARN se producen errores que no pueden ser subsanados por enzimas celulares. De esta manera se originan virus nuevos con pequeñas diferencias, mutaciones, a veces imperceptibles. Estas mezclas de virus nuevos se llaman cuasiespecies. Otras veces, las mutaciones son más importantes y dan lugar a tipos distintos para la misma especie. Así, los VP presentan tres tipos diferentes, a saber, tipo 1, 2 y 3. Estos tipos se diferencian en las proteínas de la nucleocápsida, hasta el punto en que los anticuerpos neutralizantes sintetizados frente a un determinado tipo de VP no actúan frente a los otros tipos. La variabilidad genética es un mecanismo evolutivo que los virus poseen al igual que otros organismos y que les ha permitido adaptarse a las distintas circunstancias y llegar hasta nuestros días, en este caso, para nuestra desgracia.
El VP fue el primer virus que se consiguió cultivar, en 1949, en líneas celulares. En 1980, se secuenció su genoma, siendo el primer virus animal con ARN secuenciado. También ha sido el primer virus infeccioso que se ha podido sintetizar sin la base de un cultivo celular, lo cual ocurrió en 1991. Gracias al éxito de las vacunas antipoliomielíticas y a los avances tecnológicos, el VP se puede utilizar como vector para conseguir determinadas proteínas y vacunas contra otros agentes infecciosos. Adaptabilidad y
resistencia Los VP resisten cualquier tipo de terapia hasta ahora conocida. También son resistentes a la acción de solventes o detergentes, y a los medios ácidos, y soportan temperaturas cercanas a los 50ºC. Son unos agentes infecciosos muy bien diseñados para permanecer mucho tiempo fuera de sus organismos huésped y poder sobrevivir.
|
[1] 10,000,000 nanómetros = 1 centímetro
Artículo publicado con permiso de la Dra. Pilar León Rega, otorgado a Organización Post-Polio México, quien es responsable sólamente de su publicación en este sitio. Edición por Ernestina Patiño. Notas de pie de página por Sergio Augusto Vistrain.
Fecha de publicación: 30 de mayo de 2005.
|
La organización El Síndrome Post-Polio
Testimonios
GRIS |

